

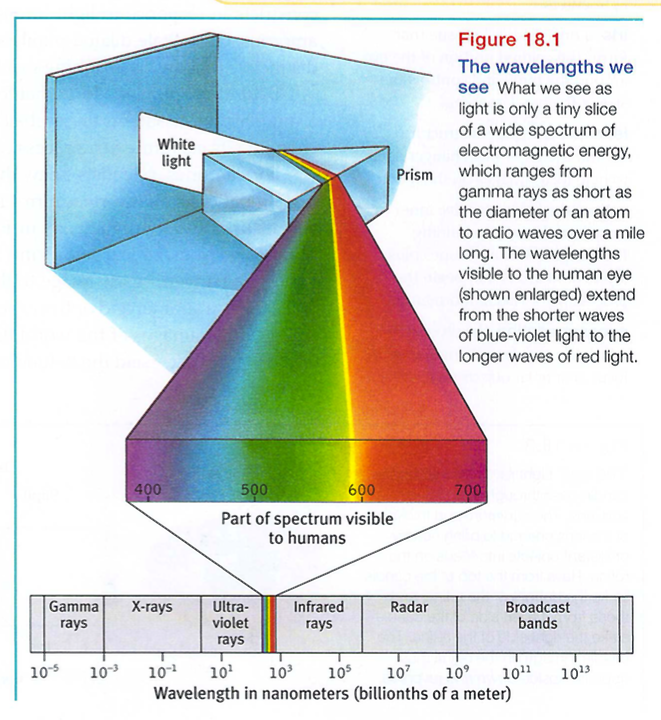
When you look at a bright red tulip, what strikes your eyes is not particles of the color red but pulses of electromagnetic energy that your visual system perceives as red. What we see as visible light is but a thin slice of the whole spectrum of electromagnetic energy, ranging from imperceptibly short gamma waves to the long waves of radio transmission (FIGURE 18.1). Other organisms are sensitive to differing portions of the spectrum. Bees, for instance, cannot see what we perceive as red but can see ultraviolet light.
Two physical characteristics of light help determine our sensory experience of them. Light's wavelength - the distance from one wave peak to the next (FIGURE 18.2a) - determines its hue (the color we experience, such as the tulip's red petals or green leaves). Intensity, the amount of energy in light waves (determined by a wave's amplitude, or height), influences brightness (Figure 18.2b). To understand how we transform physical energy into color and meaning, we first need to understand vision's window, the eye.
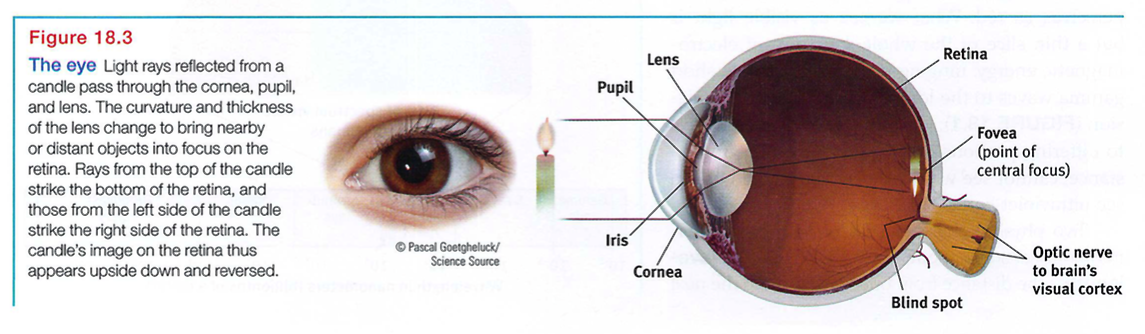
Light enters the eye through the cornea, which protects the eye and bends light to provide focus (FIGURE 18.3). The light then passes through the pupil, a small adjustable opening. Surrounding the pupil and controlling its size is the iris, a colored muscle that dilates or constricts in response to light intensity and even to inner emotions. (When we're feeling amorous, our telltale dilated pupils and dark eyes subtly signal our interest.) Each iris is so distinctive that an iris - scanning machine can confirm your identity.
Behind the pupil is a lens that focuses incoming light rays into an image on the retina, a multilayered tissue on the eyeball's sensitive inner surface. The lens focuses the rays by changing its curvature in a process called accommodation.
For centuries, scientists knew that when an image of a candle passes through a small opening, it casts an inverted mirror image on a dark wall behind. If the retina receives this sort of upside - down image, as in Figure 18.3, how can we see the world right side up? The ever - curious Leonardo da Vinci had an idea: Perhaps the eye's watery fluids bend the light rays, reinverting the image to the upright position as it reaches the retina. But then in 1604, the astronomer and optics expert Johannes Kepler showed that the retina does receive upside - down images of the world (Crombie, 1964). And how could we understand such a world? "I leave it," said the befuddled Kepler, "to natural philosophers."
Eventually, the answer became clear: The retina doesn't "see" a whole image. Rather, its millions of receptor cells convert particles of light energy into neural impulses and forward those to the brain. There, the impulses are reassembled into a perceived, upright-seeming image.
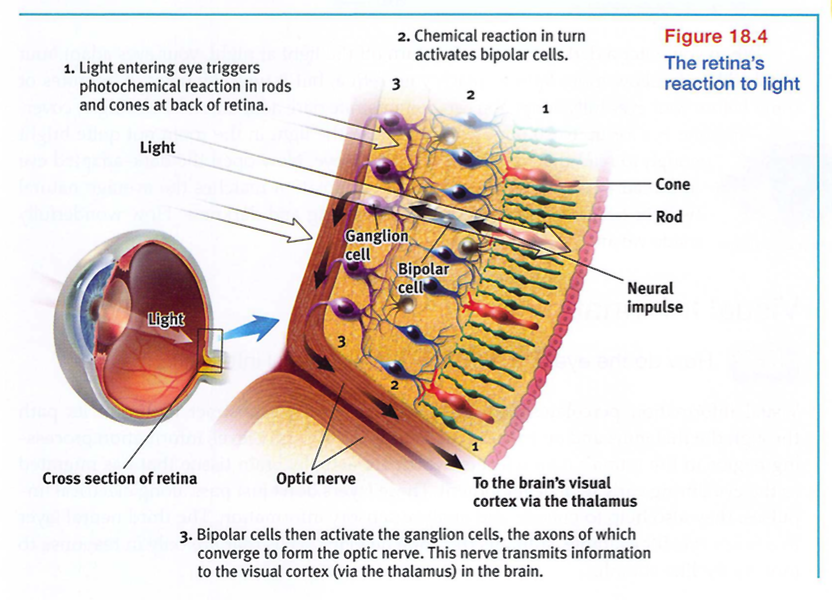
If you could follow a single light - energy particle into your eye, you would first make your way through the retina's outer layer of cells to its buried receptor cells, the rods and cones (FIGURE 18.4). There, you would see the light energy trigger chemical changes that would spark neural signals, activating nearby bipolar cells. The bipolar cells in turn would activate the neighboring ganglion cells, whose axons twine together like the strands of a rope to form the optic nerve. That nerve will carry the information to your brain, where your thalamus stands ready to distribute the information. The optic nerve can send nearly 1 million messages at once through its nearly 1 million ganglion fibers. (The auditory nerve, which enables hearing, carries much less information through its mere 30,000 fibers.) We pay a small price for this eye-to-brain highway. Where the optic nerve leaves the eye, there are no receptor cells - creating a blind spot (FIGURE 18.5 on the next page). Close one eye and you won't see a black hole, however. Without seeking your approval, your brain fills in the hole.
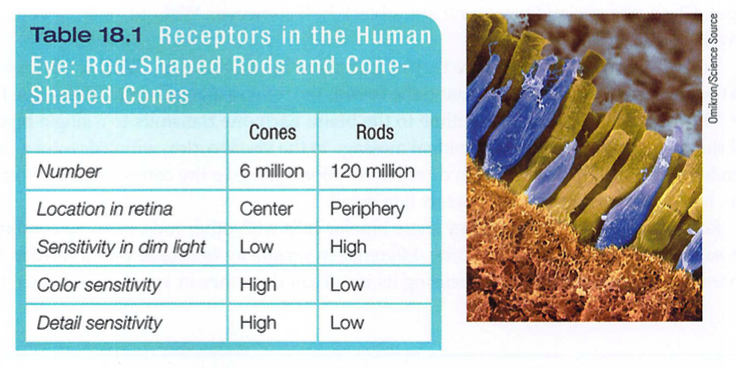
Rods and cones differ in where they're found and in what they do (TABLE 18.1). Cones cluster in and around the fovea, the retina's area of central focus (see Figure 18.3). Many have their own hotline to the brain: Each one transmits to a single bipolar cell that helps relay the cone's individual message to the visual cortex, which devotes a large area to input from the fovea. These direct connections preserve the cones' precise information' making them better able to detect fine detail.
Rods have no such hotline; they share bipolar cells with other rods, sending combined messages. To experience this rod-cone difference in sensitivity to details, pick a word in this sentence and stare directly at it, focusing its image on the cones in your fovea. Notice that words a few inches off to the side appear blurred? Their image strikes the outer regions of your retina, where rods predominate. Thus, when driving or biking, you can detect a car in your peripheral vision well before perceiving its details.
Cones also enable you to perceive color. In dim light they become ineffectual, so you see no colors. Rods, which enable black-and-white vision, remain sensitive in dim light. Several rods will funnel their faint energy output onto a single bipolar cell. Thus, cones and rods each provide a special sensitivity - cones to detail and color, and rods to faint light.
When you enter a darkened theater or turn off the light at night, your eyes adapt. Your pupils dilate to allow more light to reach your retina, but it typically takes 20 minutes or more before your eyes fully adapt. You can demonstrate dark adaptation by closing or covering one eye for up to 20 minutes. Then make the light in the room not quite bright enough to read this book with your open eye. Now open the dark-adapted eye and read (easily). This period of dark adaptation matches the average natural twilight transition between the Sun's setting and darkness. How wonderfully made we are.
How do the eye and the brain process visual information?
Visual information percolates through progressively more abstract levels on its path through the thalamus and on to the visual cortex. At the entry level, information processing begins in the retina's neural layers, which are actually brain tissue that has migrated to the eye during early fetal development. These layers don't just pass along electrical impulses; they also help to encode and analyze sensory information. The third neural layer in a frog's eye, for example, contains the "bug detector" cells that fire only in response to moving fly - like stimuli.
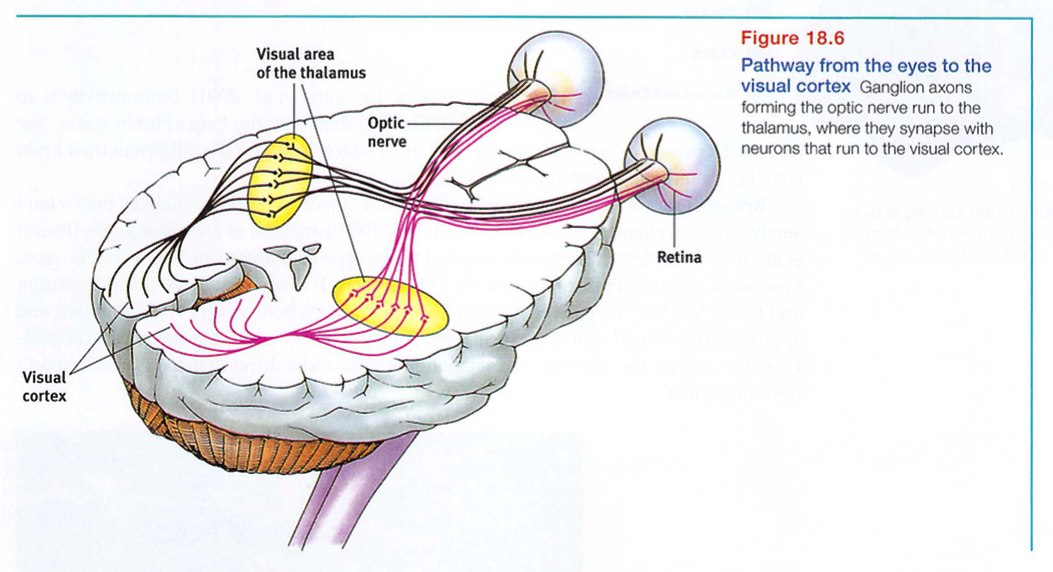
After processing by your retina's nearly 130 million receptor rods and cones, information travels to your bipolar cells, then to your million or so ganglion cells, and through their axons making up the optic nerve to your brain. Any given retinal area relays its information to a corresponding location in the visual cortex, in the occipital lobe at the back of your brain (FIGURE 18.6).
The same sensitivity that enables retinal cells to fire messages can lead them to misfire, as you can demonstrate for yourself. Turn your eyes to the left, close them, and then gently rub the right side of your right eyelid with your fingertip. Note the patch of light to the left, moving as your finger moves. Why do you see light? Why at the left?
Your retinal cells are so responsive that even pressure triggers them. But your brain interprets their firing as light. Moreover, it interprets the light as coming from the left - the normal direction of light that activates the right side of the retina.
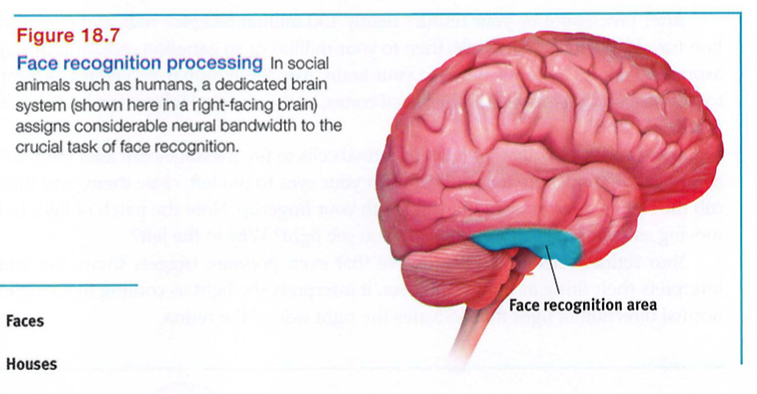
David Hubel and Torsten Wiesel (1979) received a Nobel Prize for their work on feature detectors. These specialized neurons in the occipital lobe's visual cortex receive information from individual ganglion cells in the retina. Feature detector cells derive their name from their ability to respond to a scene's specific features - to particular edges, lines, angles, and movements. These cells pass this information to other cortical areas, where teams of cells (supercell clusters) respond to more complex patterns. As we noted in Module 12, one temporal lobe area by your right ear (FIGURE 18.7) enables you to perceive faces and, thanks to a specialized neural network, to recognize them from varied viewpoints (Connor, 2010). If this region were damaged, you might recognize other forms and objects, but, like Heather Sellers, not familiar faces. When researchers temporarily disrupt the brain's face - processing areas with magnetic pulses, people are unable to recognize faces .
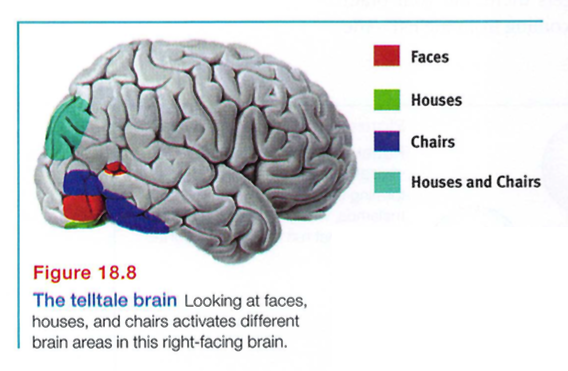
They will, however, be able to recognize houses, because the brain's face - perception occurs separately from its object - perception (McKone et al., 2007; Pitcher et al., 2007). Thus, functional MRI (fMRI) scans show different brain areas activating when people view varied objects (Downing et a1., 2001). Brain activity is so specific (FIGURE 18.8) that, with the help of brain scans, "we can tell if a person is looking at a shoe, a chair, or a face, based on the pattern of their brain activity," noted one researcher (Haxby, 2001).
Research shows that for biologically important objects and events, monkey brains (and surely ours as well) have a "vast visual encyclopedia" distributed as specialized cells (Perrett et a1., 1988, 1992, 1994). These cells respond to one type of stimulus, such as a specific gaze, head angle, posture, or body movement. Other supercell clusters integrate this information and fire only when the cues collectively indicate the direction of someone's attention and approach. This instant analysis, which aided our ancestors' survival, also helps a soccer goalkeeper anticipate the direction of an impending kick, and a driver anticipate a pedestrian's next movement.
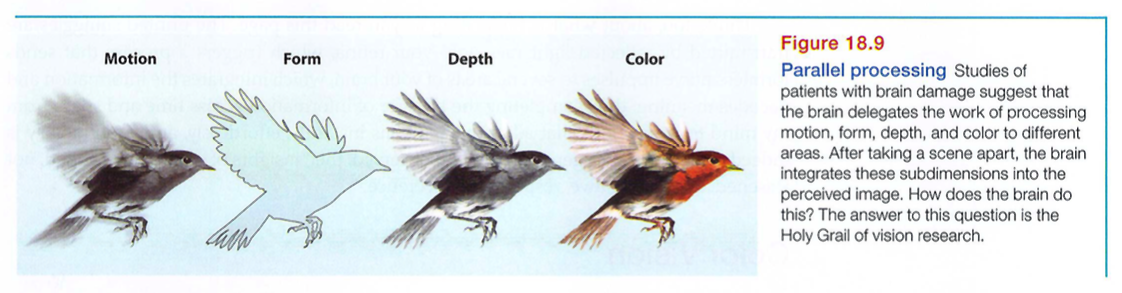
Our brain achieves these and other remarkable feats by means of parallel processing: doing many things at once. To analyze a visual scene, the brain divides it into subdimensions - motion, form, depth, color - and works on each aspect simultaneously (Livingstone & Hubel, 1988). We then construct our perceptions by integrating the separate but parallel work of these different visual teams (FIGURE 18.9).
To recognize a face, your brain integrates information projected by your retinas to several visual cortex areas, compares it with stored information, and enables you to recognize the face: Grandmother! Scientists are debating whether this stored information is contained in a single cell or distributed over a network. Some supercells - "grandmother cells" - do appear to respond very selectively to 1 or 2 faces in 100 (Bowers, 2009) . The whole facial recognition process requires tremendous brain power - 30 percent of the cortex (10 times the brain area devoted to hearing).
Destroy or disable a neural workstation for a visual sub task, and something peculiar results, as happened to "Mrs. M." (Hoffman, 1998). Since a stroke damaged areas near the rear of both sides of her brain, she has been unable to perceive movement. People in a room seem "suddenly here or there but I have not seen them moving." Pouring tea into a cup is a challenge because the fluid appears frozen - she cannot perceive it rising in the cup.
After stroke or surgery damage to the brain's visual cortex, others have experienced blind-sight (a phenomenon we met in Module 13). Shown a series of sticks, they report seeing nothing. Yet when asked to guess whether the sticks are vertical or horizontal, their visual intuition typically offers the correct response. When told, "You got them all right," they are astounded. There is, it seems, a second"mind" - a parallel processing system - operating unseen. These separate visual systems for perception and action illustrate dual processing - the two-track mind.
* * *
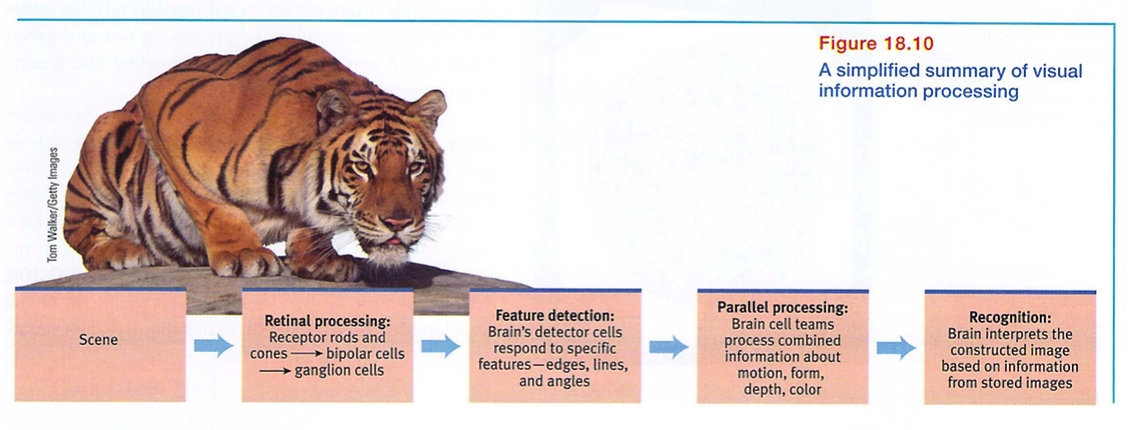
Think about the wonders of visual processing. As you look at that tiger in the zoo, information enters your eyes, is transduced, and is sent to your brain as millions of neural impulses. As your brain buzzes with activity, various areas focus on different aspects of the tiger's image. Finally, in some as yet mysterious way, these separate teams pool their work to produce a meaningful image, which you compare with previously stored images and recognize: a crouching tiger (FIGURE 18.10).
Think, too, about what is happening as you read this page. The printed squiggles are transmitted by reflected light rays onto your retina, which triggers a process that sends formless nerve impulses to several areas of your brain, which integrates the information and decodes meaning, thus completing the transfer of information across time and space from my mind to your mind. That all of this happens instantly, effortlessly, and continuously is indeed awesome. As Roger Sperry (1985) observed, the "insights of science give added, not lessened, reasons for awe, respect, and reverence."
What theories help us understand color vision?
We talk as though objects possess color: "A tomato is red." Perhaps you have pondered the old question, "If a tree falls in the forest and no one hears it, does it make a sound?" We can ask the same of color: If no one sees the tomato, is it red?
The answer is No. First, the tomato is everything but red, because it rejects (reflects) the long wavelengths of red. Second, the tomato's color is our mental construction. As Isaac Newton (1704) noted, "The [light] rays are not colored." Color, like all aspects of vision, resides not in the object but in the theater of our brain, as evidenced by our dreaming in color.
One of vision's most basic and intriguing mysteries is how we see the world in color. How, from the light energy striking the retina, does the brain manufacture our experience of color - and of such a multitude of colors? Our difference threshold for colors is so low that we can discriminate more than 1 million different color variations (Neitz et a1., 2001) . At least most of us can. For about 1 person in 50, vision is color deficient - and that person is usually male, because the defect is genetically sex - linked.
Why is some people's vision deficient? To answer that question, we need to understand how normal color vision works. Modern detective work on this mystery began in the nineteenth century, when Hermann von Helmholtz built on the insights of an English physicist, Thomas Young. Knowing that any color can be created by combining the light waves of three primary colors - red, green, and blue - Young and von Helmholtz inferred that the eye must have three corresponding types of color receptors. Years later, researchers measured the response of various cones to different color stimuli and confirmed the Young - Helmholtz trichromatic (three - color) theory, which implies that the receptors do their color magic in teams of three. Indeed, the retina has three types of color receptors, each especially sensitive to one of three colors. And those colors are, in fact, red, green, and blue. When we stimulate combinations of these cones, we see other colors. For example, there are no receptors especially sensitive to yellow. We see yellow when mixing red and green light, which stimulates both red-sensitive and green-sensitive cones. Most people with color-deficient vision are not actually "colorblind." They simply lack functioning red- or green-ensitive cones, or sometimes both. Their vision - perhaps unknown to them, because their lifelong vision seems normal - is monochromatic (one - color) or dichromatic (two - color) instead of trichromatic, making it impossible to distinguish the red and green in FIGURE 18.11 (Boynton, 1979). Dogs, too, lack receptors for the wavelengths of red, giving them only limited, dichromatic color vision (Neitz et al., 1989).
But how is it that people blind to red and green can often still see yellow? And why does yellow appear to be a pure color and not a mixture of red and green, the way purple is of red and blue? As Ewald Hering soon noted, trichromatic theory leaves some parts of the color vision mystery unsolved.
Hering, a physiologist, found a clue in afterimages. Stare at a green square for a while and then look at a white sheet of paper, and you will see red, green's opponent color. Stare at a yellow square and its opponent color, blue, will appear on the white paper. (To experience this, try the flag demonstration in FIGURE 18.12.) Hering surmised that there must be two additional color processes, one responsible for red-versus-green perception, and one for blue-versus-yellow.
Indeed, a century later, researchers also confirmed Hering's opponent-process theory. Three sets of opponent retinal processes - red-green, yellow-blue, and white-black - enable color vision. In the retina and in the thalamus (where impulses from the retina are relayed en route to the visual cortex), some neurons are turned "on" by red but turned "off" by green. Others are turned on by green but off by red (DeValois & DeValois, 1975). Like red and green marbles sent down a narrow tube, "red" and "green" messages cannot both travel at once. So we do not experience a reddish green. (Red and green are thus opponents.) But red and blue travel in separate channels, so we can see a reddish-blue magenta.
So how do we explain afterimages, such as in the flag demonstration? By staring at green, we tire our green response. When we then stare at white (which contains all colors, including red), only the red part of the green - red pairing will fire normally.
The present solution to the mystery of color vision is therefore roughly this: Color processing occurs in two stages. The retina's red, green, and blue cones respond in varying degrees to different color stimuli, as the Young - Helmholtz trichromatic theory suggested. Their signals are then processed by the nervous system's opponent - process cells, as Hering's theory proposed.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}