

In Arthur Conan Doyle's A Study in Scarlet, Sherlock Holmes offers a popular theory of memory capacity:
I consider that a man's brain originally is like a little empty attic, and you have to stock it with such furniture as you choose....It is a mistake to think that that little room has elastic walls and can distend to any extent. Depend upon it, there comes a time when for every addition of knowledge you forget something that you knew before.
Contrary to Holmes' "memory model," our capacity for storing long-term memories is essentially limitless. Our brains are not like attics, which once filled can store more items only if we discard old ones.
I marveled at my aging mother-in-law, a retired pianist and organist. At age 88, her blind eyes could no longer read music. But let her sit at a keyboard and she would flawlessly play any of hundreds of hymns, including ones she had not thought of for 20 years. Where did her brain store those thousands of sequenced notes?
For a time, some surgeons and memory researchers marveled at patients' seeming vivid memories triggered by brain stimulation during surgery. Did this prove that our whole past, not just well-practiced music, is"in there,"in complete detail, just waiting to be relived? On closer analysis, the seeming flashbacks appeared to have been invented, not relived (Loftus & Loftus, 1980). In a further demonstration that memories do not reside in single, specific spots, psychologist Karl Lashley (1950) trained rats to find their way out of a maze, then surgically re¬moved pieces of their brain's cortex and retested their memory. No matter which small brain section he removed, the rats retained at least a partial memory of how to navigate the maze.
The point to remember: Despite the brain's vast storage capacity, we do not store information as libraries store their books, in discrete, precise locations. Instead, many parts of the brain interact as we encode, store, and retrieve the information that forms our memories.
As with perception, language, emotion, and much more, memory requires brain networks. The network that processes and stores your explicit memories for facts and episodes includes your frontal lobes and hippocampus. When you summon up a mental encore of a past experience, many brain regions send input to your frontal lobes for working memory processing (Fink et al., 1996; Gabrieli et al., 1996; Markowitsch, 1995). The left and right frontal lobes process different types of memories. Recalling a password and holding it in working memory, for example, would activate the left frontal lobe. Calling up a visual party scene would more likely activate the right frontal lobe.
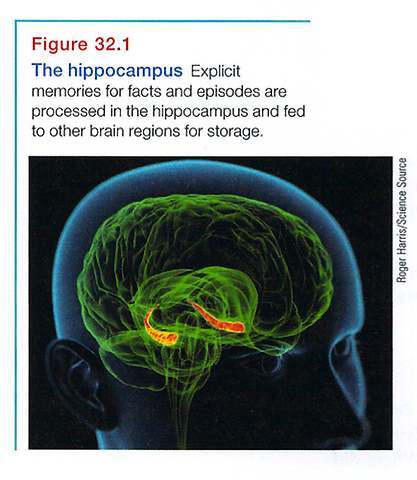
Cognitive neuroscientists have found that the hippocampus, a temporal-lobe neural center located in the limbic system, is the brain's equivalent of a"save" button for explicit memories (see FIGURE 32.1). Brain scans, such as PET scans of people recalling words, and autopsies of people who had amnesia (memory loss) have revealed that new explicit memories of names, images, and events are laid down via the hippocampus (Squire, 1992).
Damage to this structure therefore disrupts recall of explicit memories. Chickadees and other birds can store food in hundreds of places and return to these unmarked caches months later-but not if their hippocampus has been removed (Kamil & Cheng, 2001; Sherry & Vaccarino, 1989). With left-hippocampus damage, people have trouble remembering verbal information, but they have no trouble recalling visual designs and locations. With right hippocampus damage, the problem is reversed (Schacter, 1996).
Subregions of the hippocampus also serve different functions. One part is active as people learn to associate names with faces (Zeineh et al., 2003). Another part is active as memory champions engage in spatial mnemonics (Maguire et al., 2003b). The rear area, which processes spatial memory, grows bigger the longer a London cabbie has navigated the maze of streets (Maguire et al., 2003a).
Memories are not permanently stored in the hippocampus. Instead, this structure seems to act as a loading dock where the brain registers and temporarily holds the elements of a remembered episode-its smell, feel, sound, and location. Then, like older files shifted to a basement storeroom, memories migrate for storage elsewhere.
Sleep supports memory consolidation. During deep sleep, the hippocampus processes memories for later retrieval. After a training experience, the greater the hippocampus activity during sleep, the better the next day's memory will be (Peigneux et a1., 2004) . Researchers have watched the hippocampus and brain cortex displaying simultaneous activity rhythms during sleep, as if they were having a dialogue (Euston et a1., 2007; Mehta, 2007). They suspect that the brain is replaying the day's experiences as it transfers them to the cortex for long-term storage. Cortex areas surrounding the hippocampus support the processing and storing of explicit memories (Squire & Zola-Morgan, 1991).
Your hippocampus and frontal lobes are processing sites for your explicit memories. But you could lose those areas and still, thanks to automatic processing, lay down implicit memories for skills and conditioned associations. Joseph LeDoux (1996) recounted the story of a brain-damaged patient whose amnesia left her unable to recognize her physician as, each day, he shook her hand and introduced himself. One day, she yanked her hand back, for the physician had pricked her with a tack in his palm. The next time he returned to introduce himself she refused to shake his hand but couldn't explain why. Having been classically conditioned, she just wouldn't do it.
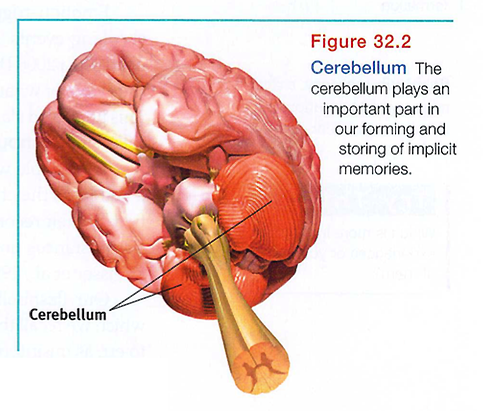
The cerebellum plays a key role in forming and storing the implicit memories created by classical conditioning. With a damaged cerebellum, people cannot develop certain conditioned reflexes, such as associating a tone with an impending puff of air-and thus do not blink in anticipation of the puff (Daum & Schugens, 1996; Green & Woodruff-Pak, 2000). When researchers surgically disrupted the function of different pathways in the cerebellum of rabbits, the rabbits became unable to learn a conditioned eye blink response (Krupa et a1., 1993; Steinmetz, 1999). Implicit memory formation needs the cerebellum (see FIGURE 32.2).
The basal ganglia, deep brain structures involved in motor movement, facilitate formation of our procedural memories for skills (Mishkin, 1982; Mishkin et al., 1997). The basal ganglia receive input from the cortex but do not return the favor of sending information back to the cortex for conscious awareness of procedural learning. If you have learned how to ride a bike, thank your basal ganglia.
Our implicit memory system, enabled partly by the cerebellum and basal ganglia, helps explain why the reactions and skills we learned during infancy reach far into our future. Yet as adults, our conscious memory of our first three years is blank, an experience called infantile amnesia. In one study, events children experienced and discussed with their mothers at age 3 were 60 percent remembered at age 7 but only 34 percent remembered at age 9 (Bauer et al., 2007). Two influences contribute to infantile amnesia: First, we index much of our explicit memory using words that nonspeaking children have not learned. Second, the hippocampus is one of the last brain structures to mature.
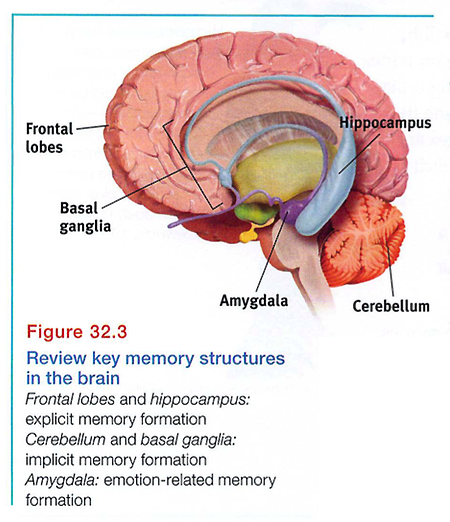
Our emotions trigger stress hormones that influence memory formation. When we are excited or stressed, these hormones make more glucose energy available to fuel brain activity, signaling the brain that something important has happened. Moreover, stress hormones provoke the amygdala (two limbic system, emotion-processing clusters) to initiate a memory trace in the frontal lobes and basal ganglia and to boost activity in the brain's memory-forming areas (Buchanan, 2007; Kensinger, 2007) (see FIGURE 32.3). The result? Emotional arousal can sear certain events into the brain, while disrupting memory for neutral events around the same time (Birnbaum et al., 2004; Brewin et al., 2007).
Emotions often persist without our conscious awareness of what caused them. In one ingenious experiment, patients with hippocampal damage (which left them unable to form new explicit memories) watched a sad film and later a happy film. After the viewing, they did not consciously recall the films, but the sad or happy emotion persisted (Feinstein et a1., 2010).
Significantly stressful events can form almost indelible (unforgettable) memories. After traumatic experiences - a school shooting, a house fire, a rape - vivid recollections of the horrific event may intrude again and again. It is as if they were burned in: " Stronger emotional experiences make for stronger, more reliable memories," noted James McGaugh (1994, 2003). This makes adaptive sense. Memory serves to predict the future and to alert us to potential dangers. Conversely, weaker emotions mean weaker memories. People given a drug that blocked the effects of stress hormones later had more trouble remembering the details of an upsetting story (Cahill, 1994).
Emotion-triggered hormonal changes help explain why we long remember exciting or shocking events, such as our first kiss or our whereabouts when learning of a loved one's death. In a 2006 Pew survey, 95 percent of American adults said they could recall exactly where they were or what they were doing when they first heard the news of the 9/11 terrorist attacks. This perceived clarity of memories of surprising, significant events leads some psychologists to call them flashbulb memories. It's as if the brain commands, "Capture this!"
The people who experienced a 1989 San Francisco earthquake did just that. A year and a half later, they had perfect recall of where they had been and what they were doing (verified by their recorded thoughts within a day or two of the quake). Others' memories for the circumstances under which they merely heard about the quake were more prone to errors (Neisser et a1., 1991; Palmer et al., 1991).
Our flashbulb memories are noteworthy for their vividness and the confidence with which we recall them. But as we relive, rehearse, and discuss them, these memories may come to err, as misinformation seeps in (Conway et a1., 2009; Talarico & Rubin, 2003, 2007).
As you read this module and think and learn about memory characteristics and processes, your brain is changing. Given increased activity in particular pathways, neural interconnections are forming and strengthening.
The quest to understand the physical basis of memory-how information becomes embedded in brain matter-has sparked study of the synaptic meeting places where neurons communicate with one another via their neurotransmitter messengers. Eric Kandel and James Schwartz (1982) observed synaptic changes during learning in the sending neurons of the California sea slug, Aplysia, a simple animal with a mere 20,000 or so unusually large and accessible nerve cells. Module 26 noted how the sea slug can be classically conditioned (with electric shock) to reflexively withdraw its gills when squirted with water, much as a shell-shocked soldier jumps at the sound of a snapping twig. By observing the slug's neural connections before and after conditioning, Kandel and Schwartz pinpointed changes. When learning occurs, the slug releases more of the neurotransmitter serotonin into certain synapses. Those synapses then become more efficient at transmitting signals.
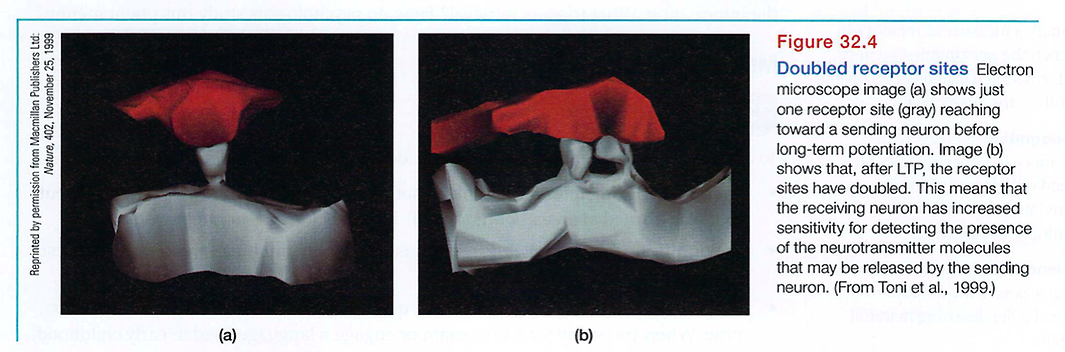
In experiments with people, rapidly stimulating certain memory-circuit connections has increased their sensitivity for hours or even weeks to come. The sending neuron now needs less prompting to release its neurotransmitter, and more connections exist between neurons (see FIGURE 32.4). This increased efficiency of potential neural firing, called longterm potentiation (LTP), provides a neural basis for learning and remembering associations (Lynch, 2002; Whitlock et al., 2006). Several lines of evidence confirm that LIP is a physical basis for memory:
Some memory-biology explorers have helped found companies that are competing to develop memory-altering drugs. The target market for memory-boosting drugs includes millions of people with Alzheimer's disease, millions more with mild neurocognitive disorder that often becomes Alzheimer's, and countless millions who would love to turn back the clock on age-related memory decline. From expanding memories perhaps will come bulging profits.
In your lifetime, will you have access to safe and legal drugs that boost your fading memory without nasty side effects and without cluttering your mind with trivia best forgotten? That question has yet to be answered. But in the meantime, one safe and free memory enhancer is already available for high schoolers everywhere: effective study techniques followed by adequate sleep! (You'll find study tips in Module 2 and at the end of this module, and sleep coverage in Modules 23 and 24.)
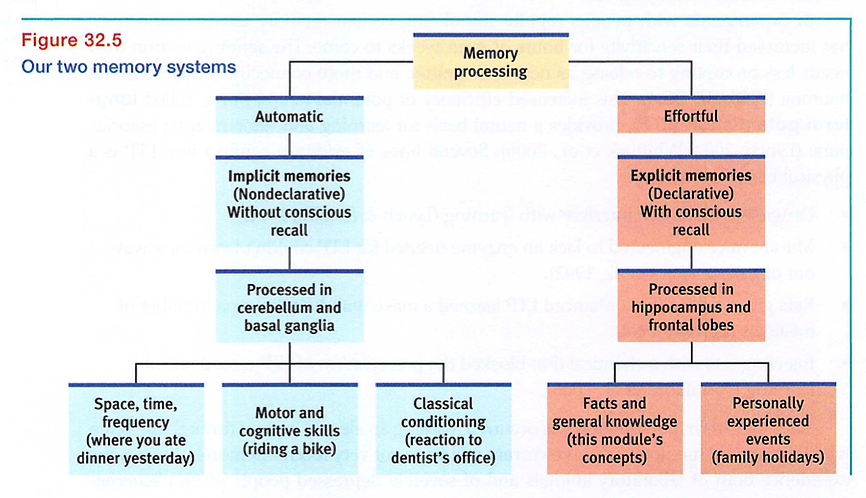
FIGURE 32.5 summarizes the brain's two-track memory processing and storage system for implicit (automatic) and explicit (effortful) memories.
After the magic of brain encoding and storage, we still have the daunting task of retrieving the information. What triggers retrieval? How do psychologists study this phenomenon?
To a psychologist, evidence of memory includes these three measures of retention:
Long after you cannot recall most of the people in your high school graduating class, you may still be able to recognize their yearbook pictures from a photographic lineup and pick their names from a list of names. In one experiment, people who had graduated 25 years earlier could not recall many of their old classmates, but they could recognize 90 percent of their pictures and names (Bahrick et al., 1975). If you are like most students, you, too, could probably recognize more names of Snow White's Seven Dwarfs than you could recall (Miserandino, 1991).
Our recognition memory is impressively quick and vast. "Is your friend wearing a new or old outfit?" "Old." "Is this 5-second movie clip from a film you've ever seen?" "Yes." "Have you ever seen this person before-this minor variation on the same old human features (two eyes, one nose, and so on)?""No." Before the mouth can form our answer to any of millions of such questions, the mind knows, and knows that it knows.
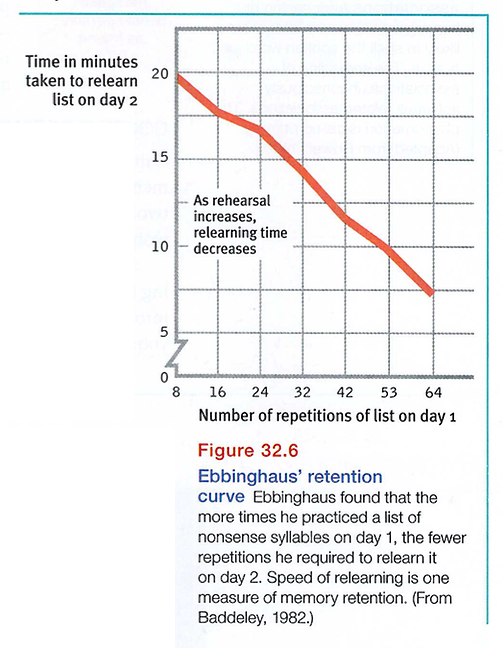
Our speed at relearning also reveals memory. Hermann Ebbinghaus showed this more than a century ago, in his learning experiments, using nonsense syllables. He randomly selected a sample of syllables, practiced them, and tested himself. To get a feel for his experiments, rapidly read aloud, eight times over, the following list (from Baddeley, 1982), then look away and try to recall the items:
JIH, BAZ, FUB,YOX, SUJ, XIR, DAX, LEQ, VUM, PID, KEL, WAV, TUV, ZOF, GEK, HIW.
The day after learning such a list, Ebbinghaus could recall few of the syllables. But they weren't entirely forgotten. As FIGURE 32.6 portrays, the more frequently he repeated the list aloud on day 1, the fewer repetitions he required to relearn the list on day 2. Additional rehearsal (overlearning) of verbal information increases retention, especially when practice is distributed over time. For students, this means that it is important to continue to rehearse course material even after you know it.
The point to remember: Tests of recognition and of time spent relearning demonstrate that we remember more than we can recall.
Imagine a spider suspended in the middle of her web, held up by the many strands extending outward from her in all directions to different points. If you were to trace a pathway to the spider, you would first need to create a path from one of these anchor points and then follow the strand down into the web.
The process of retrieving a memory follows a similar principle, because memories are held in storage by a web of associations, each piece of information interconnected with others. When you encode into memory a target piece of information, such as the name of the person sitting next to you in class, you associate with it other bits of information about your surroundings, mood, seating position, and so on. These bits can serve as retrieval cues that you can later use to access the information. The more retrieval cues you have, the better your chances of finding a route to the suspended memory.
The best retrieval cues come from associations we form at the time we encode a memory - smells, tastes, and sights that can evoke our memory of the associated person or event. To call up visual cues when trying to recall something, we may mentally place ourselves in the original context. After losing his sight, British scholar John Hull (1990, p. 174) described his difficulty recalling such details:
I knew I had been somewhere, and had done particular things with certain people, but where? I could not put the conversations ... into a context. There was no background, no features against which to identify the place. Normally, the memories of people you have spoken to during the day are stored in frames which include the background.
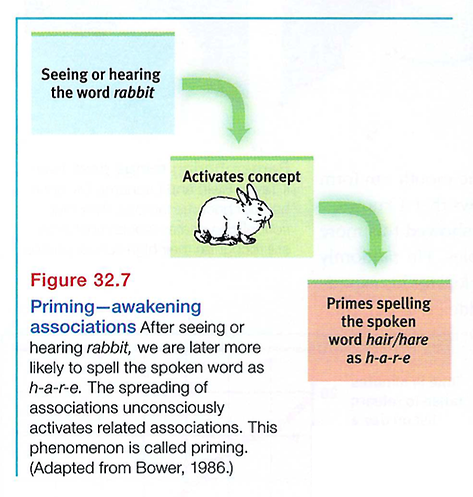
Often our associations are activated without our awareness. The philosopher-psychologist William James referred to this process, which we call priming, as the "wakening of associations." Seeing or hearing the word rabbit primes associations with hare, even though we may not recall having seen or heard rabbit (see FIGURE 32.7).
Priming is often “memoryless memory" -invisible memory, without your conscious awareness. If, walking down a hallway, you see a poster of a missing child, you may then unconsciously be primed to interpret an ambiguous adult-child interaction as a possible kidnapping Games, 1986). Although you no longer have the poster in mind, it predisposes your interpretation.
Priming can influence behaviors as well. In one study, participants primed with money-related words were less likely to help another person when asked (Vohs et al., 2006). In such cases, money may prime our materialism and self-interest rather than the social norms that encourage us to help (Ariely, 2009).
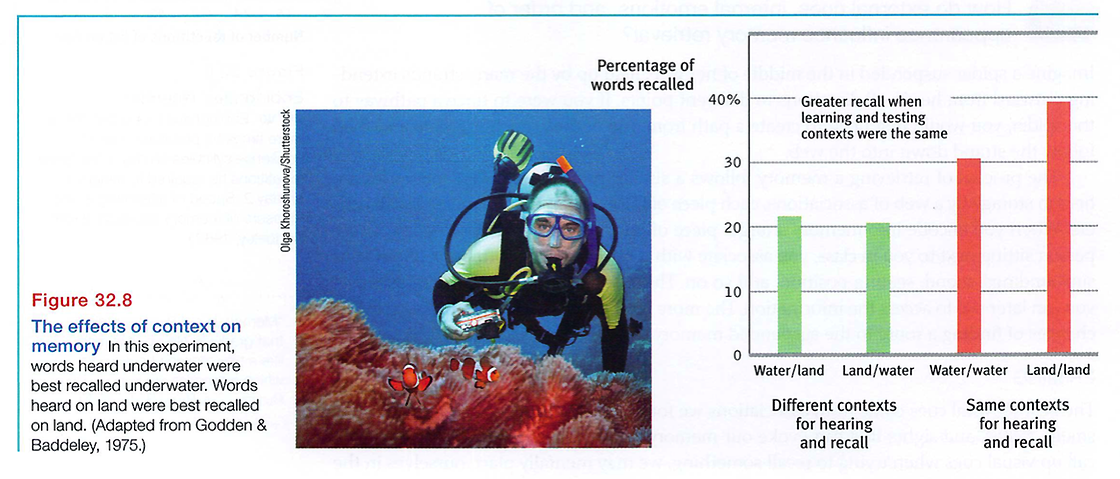
Putting yourself back in the context where you experienced something can prime your memory retrieval. As FIGURE 32.8 illustrates, when scuba divers listened to a word list in two different settings (either 10 feet underwater or sitting on the beach), they recalled more words if retested in the same place (Godden & Baddeley, 1975).
You may have experienced similar context effects. Consider this scenario: While taking notes from this book, you realize you need to sharpen your pencil. You get up and walk into another room, but then you cannot remember why. After returning to your desk it hits you: "I wanted to sharpen this pencil!" What happens to create this frustrating experience? In one context (desk, reading psychology), you realize your pencil needs sharpening. When you go to the other room and are in a different context, you have few cues to lead you back to that thought. When you are once again at your desk, you are back in the context in which you encoded the thought ("This pencil is dull").
In several experiments, one researcher found that a familiar context could activate memories even in 3-month-olds (Rovee-Collier, 1993). After infants learned that kicking a crib mobile would make it move (via a connecting ribbon from the ankle), the infants kicked more when tested again in the same crib with the same bumper than when in a different context.
Closely related to context-dependent memory is state-dependent memory. What we learn in one state-be it drunk or sober-may be more easily recalled when we are again in that state. What people learn when drink they don't recall well in any state (alcohol disrupts storage). But they recall it slightly better when again drunk. Someone who hides money when drunk may forget the location until drunk again.
Our mood states provide an example of memory's state dependence. Emotions that accompany good or bad events become retrieval cues (Fiedler et a1., 2001). Thus, our memories are somewhat mood congruent. If you've had a bad evening-your date never showed, your Chicago Cubs hat disappeared, your TV went out 10 minutes before the end of a show-your gloomy mood may facilitate recalling other bad times. Being depressed sours memories by priming negative associations, which we then use to ex plain our current mood. In many experiments, people put in a buoyant mood - whether under hypnosis or just by the day's events (a World Cup soccer victory for German participants in one study) - have recalled the world through rose-colored glasses (DeSteno et al., 2000; Forgas et al., 1984; Schwarz et a1., 1987). They judged themselves competent and effective, other people benevolent, happy events more likely.
Knowing this mood-memory connection, we should not be surprised that in some studies currently depressed people have recalled their parents as rejecting, punitive, and guilt promoting, whereas formerly depressed people's recollections more closely resembled the more positive descriptions given by those who never suffered depression (Lewinsohn & Rosenbaum, 1987; Lewis, 1992). Similarly, adolescents'ratings of parental warmth in one week gave little clue to how they would rate their parents six weeks later (Bornstein et a1., 1991). When teens were down, their parents seemed inhuman; as their mood brightened, their parents morphed from devils into angels. In a good or bad mood, we persist in attributing to reality our own changing judgments, memories, and interpretations. In a bad mood, we may read someone's look as a glare and feel even worse. In a good mood, we may encode the same look as interest and feel even better. Passions exaggerate.
This retrieval effect helps explain why our moods persist. When happy, we recall happy events and therefore see the world as a happy place, which helps prolong our good mood. When depressed, we recall sad events, which darkens our interpretations of current events. For those of us with a predisposition to depression, this process can help maintain a vicious, dark cycle.
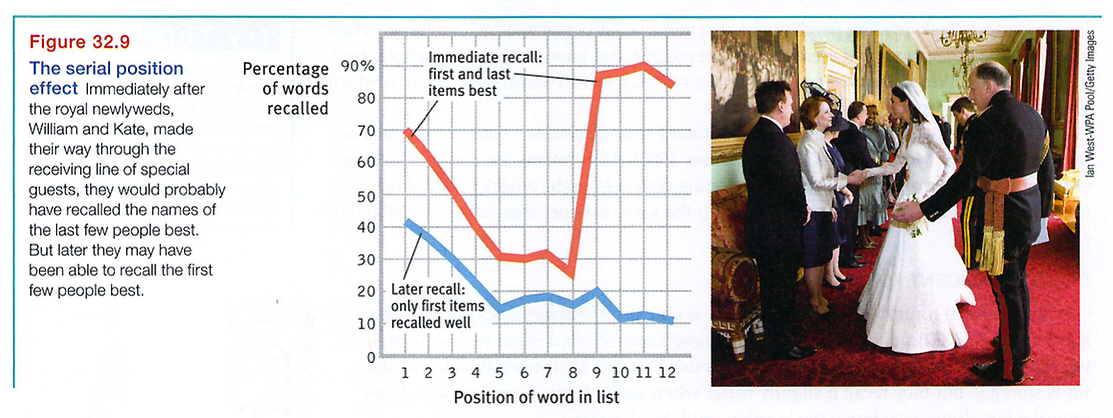
Another memory-retrieval quirk, the serial position effect, can leave us wondering why we have large holes in our memory of a list of recent events. Imagine it's your first day in a new job, and your manager is introducing co-workers. As you meet each person, you silently repeat everyone's name, starting from the beginning. As the last person smiles and turns away, you feel confident you'll be able to greet your new co-workers by name the next day.
Don't count on it. Because you have spent more time rehearsing the earlier names than the later ones, those are the names you'll probably recall more easily the next day.
In experiments, when people view a list of items (words, names, dates, even odors) and immediately try to recall them in any order, they fall prey to the serial position effect (Reed, 2000). They briefly recall the last items especially quickly and well (a recency effect), perhaps because those last items are still in working memory. But after a delay, when they have shifted their attention away from the last items, their recall is best for the first items (a primacy effect; see FIGURE 32.9).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}